Eukaryotic Translation Initiation Factor 4E Antibodies
Your search for reliable EIF4E antibodies ends here. EIF4E, known by aliases such as EIF4E, eif4ea, Eif4e, eIF-4E, eif6, eif4e, eif4e.L, eif4e1b.S, LOC100533326, eif4eb, is an integral part of our antibody range. Whether you're working with Human, Rat, Mouse, Zebrafish (Danio rerio), Cow, or other species, our range of EIF4E antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, ELISA, IHC, FACS, IF (cc), providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our EIF4E antibodies in your research endeavors for dependable EIF4E detection.
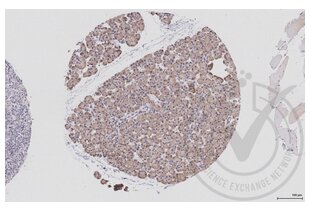
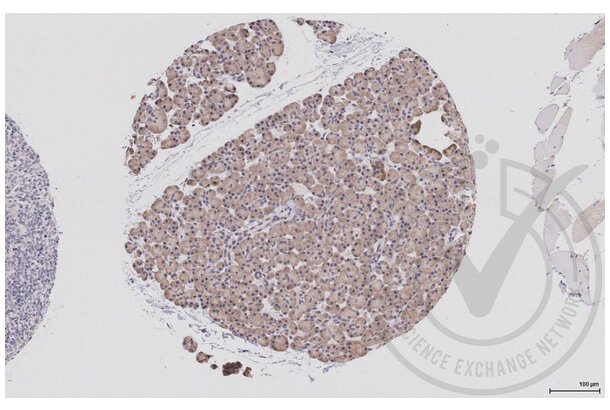 EIF4E antibody (AA 121-217) (ABIN1386772)
EIF4E antibody (AA 121-217) (ABIN1386772)
EIF4E Reactivity: Human, Rat WB, ELISA, IHC (p), IF (cc), IF (p), ICC, IHC (fro) Host: Rabbit Polyclonal unconjugated
EIF4E Reactivity: Human WB, IHC, ELISA, FACS, ICC Host: Mouse Monoclonal 5D11 unconjugated
EIF4E Reactivity: Human WB, IHC, IF, IP Host: Rabbit Polyclonal unconjugated
Eukaryotic Translation Initiation Factor 4E Antibodies by Reactivity
Find Eukaryotic Translation Initiation Factor 4E Antibodies for a variety of species such as anti-Human Eukaryotic Translation Initiation Factor 4E, anti-Rat Eukaryotic Translation Initiation Factor 4E, anti-Mouse Eukaryotic Translation Initiation Factor 4E. The species listed below are among those available. Click on a link to go to the corresponding products.
Eukaryotic Translation Initiation Factor 4E Antibodies by Application
Find Eukaryotic Translation Initiation Factor 4E Antibodies validated for a specific application such as WB, ELISA, IHC, FACS. Some of the available applications are listed below. Click on a link to go to the corresponding products.
Eukaryotic Translation Initiation Factor 4E Antibodies by Host
Find Eukaryotic Translation Initiation Factor 4E Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
Eukaryotic Translation Initiation Factor 4E Antibodies by Clonality
Find available monoclonal or polyclonal Eukaryotic Translation Initiation Factor 4E Antibodies. Click on a link to go to the corresponding products.
Eukaryotic Translation Initiation Factor 4E Antibodies by Clone
Find Eukaryotic Translation Initiation Factor 4E Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
Popular Eukaryotic Translation Initiation Factor 4E Antibodies
- (3)
- (1)
- (2)
- (6)
- (2)
- (9)
- (4)
- (4)
- (5)
- (3)
- (3)
- (1)
- (2)
- (3)
- (3)
- (4)
- (2)
- (2)
- (2)
Latest Publications for our Eukaryotic Translation Initiation Factor 4E Antibodies
: "CUL3 Deficiency Causes Social Deficits and Anxiety-like Behaviors by Impairing Excitation-Inhibition Balance through the Promotion of Cap-Dependent Translation." in: Neuron, (2019) (PubMed).: "20(s)-Protopanaxadiol (PPD) increases the radiotherapy sensitivity of laryngeal carcinoma." in: Food & function, Vol. 8, Issue 12, pp. 4469-4477, (2018) (PubMed).
: "Picornavirus 2A protease regulates stress granule formation to facilitate viral translation." in: PLoS pathogens, Vol. 14, Issue 2, pp. e1006901, (2018) (PubMed).
: "Inclusion bodies of human parainfluenza virus type 3 inhibit antiviral stress granule formation by shielding viral RNAs." in: PLoS pathogens, Vol. 14, Issue 3, pp. e1006948, (2018) (PubMed).
: "Diminished satellite cell fusion and S6K1 expression in myotubes derived from skeletal muscle of low birth weight neonatal pigs." in: Physiological reports, Vol. 5, Issue 3, (2017) (PubMed).
: "A fine balance: Regulation of hippocampal Arc/Arg3.1 transcription, translation and degradation in a rat model of normal cognitive aging." in: Neurobiology of learning and memory, Vol. 115, pp. 58-67, (2014) (PubMed).
: "Multiple components of eIF4F are required for protein synthesis-dependent hippocampal long-term potentiation." in: Journal of neurophysiology, Vol. 109, Issue 1, pp. 68-76, (2013) (PubMed).
: "Circadian proteins CLOCK and BMAL1 in the chromatoid body, a RNA processing granule of male germ cells." in: PLoS ONE, Vol. 7, Issue 8, pp. e42695, (2012) (PubMed).
: "Inhibition of the interactions between eukaryotic initiation factors 4E and 4G impairs long-term associative memory consolidation but not reconsolidation." in: Proceedings of the National Academy of Sciences of the United States of America, Vol. 108, Issue 8, pp. 3383-8, (2011) (PubMed).
: "Kinetic mechanism for assembly of the m7GpppG.eIF4E.eIF4G complex." in: The Journal of biological chemistry, Vol. 283, Issue 37, pp. 25227-37, (2008) (PubMed).
Aliases for Eukaryotic Translation Initiation Factor 4E Antibodies
eukaryotic translation initiation factor 4E (EIF4E) Antibodieseukaryotic translation initiation factor 4ea (eif4ea) Antibodies
eukaryotic translation initiation factor 4E (Eif4e) Antibodies
Eukaryotic initiation factor 4E (eIF-4E) Antibodies
eucaryotic initiation factor 6 (eif6) Antibodies
eukaryotic translation initiation factor 4E (eif4e) Antibodies
eukaryotic translation initiation factor 4E L homeolog (eif4e.L) Antibodies
eukaryotic translation initiation factor 4E family member 1B S homeolog (eif4e1b.S) Antibodies
translation initiation factor eIF4E (LOC100533326) Antibodies
eukaryotic translation initiation factor 4eb (eif4eb) Antibodies
2 Antibodies
0260/09 Antibodies
0587/11 Antibodies
0589/11 Antibodies
0919/12 Antibodies
1004/13 Antibodies
7238 Antibodies
AUTS19 Antibodies
CBP Antibodies
cbp Antibodies
CG4035 Antibodies
CT13384 Antibodies
CT39424 Antibodies
CT39426 Antibodies
D-eIF4E Antibodies
deIF-4E Antibodies
dEif4e Antibodies
deIF4E Antibodies
Dmel\\CG4035 Antibodies
EG668879 Antibodies
eIF-4E Antibodies
eIF-4e Antibodies
eif-4E Antibodies
eif-4e Antibodies
EIF-4E Antibodies
eIF-4E1 Antibodies
eIF-4E2 Antibodies
eIF-4EII Antibodies
Eif4E Antibodies
Eif4e Antibodies
eIF 4E Antibodies
eIF4E Antibodies
eIF4e Antibodies
eif4e Antibodies
eif4e-1 Antibodies
eIF4E-1 Antibodies
eIF4E-1/2 Antibodies
eIF4E-2 Antibodies
Eif4e-ps Antibodies
EIF4E1 Antibodies
eIF4E1 Antibodies
eif4e1 Antibodies
eIF4EI Antibodies
EIF4EL1 Antibodies
eif4el1 Antibodies
EIF4F Antibodies
eif4f Antibodies
If4e Antibodies
IF4E Antibodies
l(3)07238 Antibodies
l(3)67Af Antibodies
l(3)S025007 Antibodies
l(3)S026009 Antibodies
l(3)S058711 Antibodies
l(3)S091912 Antibodies
zgc:86680 Antibodies
zgc:101581 Antibodies
Did you look for something else?
- Eukaryotic Translation Initiation Factor 3, Subunit M Antibodies
- Eukaryotic Translation Initiation Factor 1B Antibodies
- Eukaryotic Translation Initiation Factor 1A Antibodies
- Eukaryotic Translation Elongation Factor 1 delta (Guanine Nucleotide Exchange Protein) Antibodies
- Eukaryotic Translation Elongation Factor 1 alpha 1 Antibodies
- ETV7 Antibodies
- ETV5 Antibodies
- ETV4 Antibodies
- ETV3L Antibodies
- ETV3 Antibodies
- ETV2 Antibodies
- ETV1 Antibodies
- ETS2 Antibodies
- Ets Variant 6 Antibodies
- ETL4 Antibodies
- ETHE1 Antibodies
- Ethanolamine Kinase 2 Antibodies
- Ethanolamine Kinase 1 Antibodies
- ETFDH Antibodies
- ETFB Antibodies
- Eukaryotic Translation Initiation Factor 4E Family Member 3 Antibodies
- Eukaryotic Translation Initiation Factor 5A Antibodies
- EVA1C Antibodies
- EVC Antibodies
- EVC2 Antibodies
- EVI2A Antibodies
- EVI2B Antibodies
- EVI5 Antibodies
- EVI5L Antibodies
- EVL Antibodies
- EVX1 Antibodies
- EVX2 Antibodies
- EWSR1 Antibodies
- EXD1 Antibodies
- EXD3 Antibodies
- Exendin 4 Antibodies
- EXO5 Antibodies
- EXOC1 Antibodies
- EXOC2 Antibodies
- EXOC3 Antibodies



