TLR9 Antibodies
Your search for reliable TLR9 antibodies ends here. TLR9, known by aliases such as TLR9, Tlr9, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Dog, Monkey, or other species, our range of TLR9 antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, ELISA, IHC, FACS, IHC (p), providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Rat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our TLR9 antibodies in your research endeavors for dependable TLR9 detection.
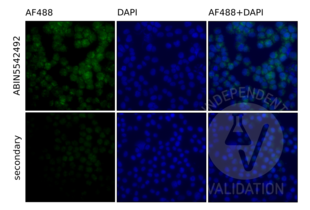
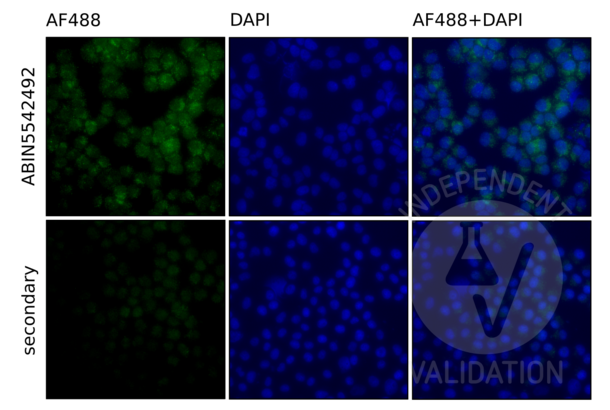 TLR9 antibody (AA 868-1016) (ABIN5542492)
TLR9 antibody (AA 868-1016) (ABIN5542492)
TLR9 Reactivity: Human IHC, ELISA, FACS, ICC Host: Mouse Monoclonal 1B12H2 unconjugated
TLR9 Reactivity: Human WB, IHC, IP, ICC Host: Rabbit Polyclonal unconjugated
TLR9 Reactivity: Human ELISA, FACS Host: Mouse Monoclonal 5B10E8 unconjugated
TLR9 Antibodies by Host
Find TLR9 Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
TLR9 Antibodies by Clonality
Find available monoclonal or polyclonal TLR9 Antibodies. Click on a link to go to the corresponding products.
TLR9 Antibodies by Conjugate
Find TLR9 Antibodies with a specific conjugate such as Biotin, FITC, PE. The conjugates listed below are among those available. Click on a link to go to the corresponding products.
Popular TLR9 Antibodies
- (7)
- (2)
- (8)
- (4)
- (4)
- (1)
- (2)
- (7)
- (2)
- (3)
- (3)
- (7)
- (2)
- (2)
- (2)
- (2)
- (2)
- (2)
- (2)
Latest Publications for our TLR9 Antibodies
: "Differential expression patterns of Toll Like Receptors and Interleukin-37 between calcific aortic and mitral valve cusps in humans." in: Cytokine, Vol. 116, pp. 150-160, (2019) (PubMed).: "Co-expression of TLR-9 and MMP-13 is associated with the degree of tumour differentiation in prostate cancer." in: International journal of experimental pathology, Vol. 100, Issue 2, pp. 123-132, (2019) (PubMed).
: "A proliferation-inducing ligand (APRIL) induced hyper-production of IgA from tonsillar mononuclear cells in patients with IgA nephropathy." in: Cellular immunology, Vol. 341, pp. 103925, (2019) (PubMed).
: "Necroptosis in pulmonary macrophages mediates lipopolysaccharide-induced lung inflammatory injury by activating ZBP-1." in: International immunopharmacology, Vol. 77, pp. 105944, (2019) (PubMed).
: "Genistein modulates the expression of Toll-like receptors in experimental autoimmune encephalomyelitis." in: Inflammation research : official journal of the European Histamine Research Society ... [et al.], Vol. 67, Issue 7, pp. 597-608, (2018) (PubMed).
: "Elevated intragraft expression of innate immunity and cell death-related markers is a risk factor for adverse graft outcome." in: Transplant immunology, Vol. 48, pp. 39-46, (2018) (PubMed).
: "High Tidal Volume Induces Mitochondria Damage and Releases Mitochondrial DNA to Aggravate the Ventilator-Induced Lung Injury." in: Frontiers in immunology, Vol. 9, pp. 1477, (2018) (PubMed).
: "Toll-like receptors 2, 4 and 9 and hypoxia markers HIF-1alpha and CAIX in pancreatic intraepithelial neoplasia." in: APMIS : acta pathologica, microbiologica, et immunologica Scandinavica, Vol. 126, Issue 11, pp. 852-863, (2018) (PubMed).
: "Impaired Surface Expression of HLA-DR, TLR2, TLR4, and TLR9 in Ex Vivo-In Vitro Stimulated Monocytes from Severely Injured Trauma Patients." in: Mediators of inflammation, Vol. 2017, pp. 2608349, (2017) (PubMed).
: "High toll-like receptor (TLR) 9 expression is associated with better prognosis in surgically treated pancreatic cancer patients." in: Virchows Archiv : an international journal of pathology, Vol. 470, Issue 4, pp. 401-410, (2017) (PubMed).
Aliases for TLR9 Antibodies
toll like receptor 9 (TLR9) Antibodiestoll-like receptor 9 (Tlr9) Antibodies
CD289 Antibodies
Did you look for something else?
- TLR8 Antibodies
- TLR7 Antibodies
- TLR6 Antibodies
- TLR5 Antibodies
- TLR4 Antibodies
- TLR3 Antibodies
- TLR2 Antibodies
- TLR13 Antibodies
- TLR12 Antibodies
- TLR11 Antibodies
- TLR10 Antibodies
- TLR1 Antibodies
- TLN2 Antibodies
- TLN1 Antibodies
- TLL1 Antibodies
- TLK2 Antibodies
- TLK1 Antibodies
- TLE6 Antibodies
- TLE4 Antibodies
- TLE3 Antibodies
- TLX1 Antibodies
- TLX2 Antibodies
- TLX3 Antibodies
- TM2D1 Antibodies
- TM2D2 Antibodies
- TM2D3 Antibodies
- TM4SF1 Antibodies
- TM4SF18 Antibodies
- TM4SF20 Antibodies
- TM4SF4 Antibodies
- TM4SF5 Antibodies
- TM6SF1 Antibodies
- TM7SF2 Antibodies
- TM9SF1 Antibodies
- TM9SF2 Antibodies
- TM9SF4 Antibodies
- TMBIM1 Antibodies
- TMBIM4 Antibodies
- TMC1 Antibodies
- Tmc2 Antibodies



