APOE ELISA Kits
Explore our high-quality APOE ELISA kits for effective APOE detection in your research. Known by various aliases including apoe, apoea, Apoe, APOE, this protein is a crucial element in our extensive ELISA kit collection. Our kits are designed for precise APOE detection across a diverse range of species, such as Human, Mouse, Rat, Dog, Rabbit, and more. They are versatile for use with various sample types, including Serum, Plasma, Tissue Homogenate, and more. The effectiveness of our kits is backed by comprehensive validations, including customer testimonials, detailed references, and illustrative images, all accessible on each product's page.
Need help finding a specific product? Our customer service team is at your service to guide you. Rely on our APOE ELISA kits for accurate and reliable APOE detection in your research projects.

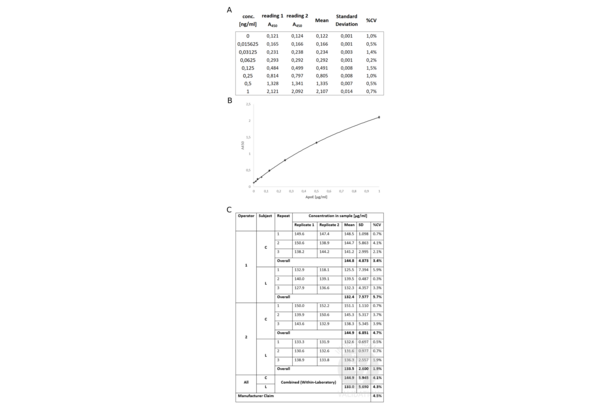 APOE ELISA Kit (ABIN612665)
APOE ELISA Kit (ABIN612665)
APOE Reactivity: Human Colorimetric Sandwich ELISA Cell Culture Supernatant, Plasma
APOE Reactivity: Human Colorimetric Sandwich ELISA 4.7 ng/mL - 300 ng/mL Cell Culture Supernatant, Cell Lysate, Plasma, Serum, Tissue Homogenate
APOE Reactivity: Mouse Colorimetric Competition ELISA 6.25 ng/mL - 400 ng/mL Cell Culture Supernatant, Plasma, Serum, Tissue Homogenate
APOE ELISA Kits by Detection Method
Find APOE ELISA Kits with a specific Detection Method. The Detection Method listed below are among those available. Click on a link to go to the corresponding products.
APOE ELISA Kits by Analytical Method
Find APOE ELISA Kits with a specific Analytical Method. The Analytical Method listed below are among those available. Click on a link to go to the corresponding products.
APOE ELISA Kits by Method Type
Find APOE ELISA Kits with a specific Method Type. The Method Type listed below are among those available. Click on a link to go to the corresponding products.
APOE ELISA Kits by Sample
Find APOE ELISA Kits with a specific Sample. The Sample listed below are among those available. Click on a link to go to the corresponding products.
APOE ELISA Kits by Binding Specificity
Find APOE ELISA Kits with a specific epitope. The epitopes listed below are among those available. Click on a link to go to the corresponding products.
Popular APOE ELISA Kits
- (1)
- (1)
- (1)
- (8)
- (1)
- (1)
- (1)
- (1)
- (1)
- (2)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
- (1)
Latest Publications for our APOE ELISA Kits
: "Hepatic HuR modulates lipid homeostasis in response to high-fat diet." in: Nature communications, Vol. 11, Issue 1, pp. 3067, (2020) (PubMed).: "Evaluation of α-synuclein and apolipoprotein E as potential biomarkers in cerebrospinal fluid to monitor pharmacotherapeutic efficacy in dopamine dictated disease states of Parkinson's disease and ..." in: Neuropsychiatric disease and treatment, Vol. 15, pp. 2073-2085, (2019) (PubMed).
: "Hypolipidemic and anti-inflammatory effects of aorta and heart tissues of cattle and pigs in the atherosclerosis rat model." in: Animal science journal = Nihon chikusan Gakkaihō, Vol. 89, Issue 5, pp. 784-793, (2018) (PubMed).
: "Deletion of plasma Phospholipid Transfer Protein (PLTP) increases microglial phagocytosis and reduces cerebral amyloid-β deposition in the J20 mouse model of Alzheimer's disease." in: Oncotarget, Vol. 9, Issue 28, pp. 19688-19703, (2018) (PubMed).
: "Serum levels of 9α,11β-PGF2 and apolipoprotein A1 achieve high predictive power as biomarkers of anaphylaxis." in: Allergy, (2017) (PubMed).
: "The involvement of ginseng berry extract in blood flow via regulation of blood coagulation in rats fed a high-fat diet." in: Journal of ginseng research, Vol. 41, Issue 2, pp. 120-126, (2017) (PubMed).
: "Adaptive immunity against gut microbiota enhances apoE-mediated immune regulation and reduces atherosclerosis and western-diet-related inflammation." in: Scientific reports, Vol. 6, pp. 29353, (2016) (PubMed).
: "A multiplex immunoassay for the non-invasive detection of bladder cancer." in: Journal of translational medicine, Vol. 14, pp. 31, (2016) (PubMed).
: "Adipocyte ATP-binding cassette G1 promotes triglyceride storage, fat mass growth, and human obesity. ..." in: Diabetes, Vol. 64, Issue 3, pp. 840-55, (2015) (PubMed).
: "Proteomic profiling of human plasma identifies apolipoprotein E as being associated with smoking and a marker for squamous metaplasia of the lung." in: Proteomics, Vol. 15, Issue 18, pp. 3267-77, (2015) (PubMed).
Aliases for APOE ELISA Kits
apolipoprotein E (apoe) ELISA Kitsapolipoprotein Ea (apoea) ELISA Kits
apolipoprotein E (Apoe) ELISA Kits
apolipoprotein E (APOE) ELISA Kits
ad2 ELISA Kits
AD2 ELISA Kits
AI255918 ELISA Kits
Apo-E ELISA Kits
apoe ELISA Kits
APOEA ELISA Kits
apoprotein ELISA Kits
im:7036787 ELISA Kits
LDLCQ5 ELISA Kits
LPG ELISA Kits
wu:fb69a05 ELISA Kits
zgc:110064 ELISA Kits
Did you look for something else?
- APOC4 ELISA Kits
- APOC3 ELISA Kits
- APOC1 ELISA Kits
- APOB ELISA Kits
- APOA5 ELISA Kits
- APOA4 ELISA Kits
- APOA2 ELISA Kits
- APOA1BP ELISA Kits
- Apo-B100 ELISA Kits
- APMAP ELISA Kits
- APLP2 ELISA Kits
- APLP1 ELISA Kits
- APITD1 ELISA Kits
- APH1A ELISA Kits
- APEX1 ELISA Kits
- Apelin Receptor ELISA Kits
- Apelin ELISA Kits
- APEH ELISA Kits
- APCS ELISA Kits
- APCDD1 ELISA Kits
- ApoE3 ELISA Kits
- APOH ELISA Kits
- APOL1 ELISA Kits
- APOLD1 ELISA Kits
- Apolipoprotein A ELISA Kits
- Apolipoprotein A-I ELISA Kits
- Apolipoprotein C-II ELISA Kits
- Apolipoprotein D ELISA Kits
- Apolipoprotein F ELISA Kits
- Apolipoprotein L 2 ELISA Kits
- Apolipoprotein M ELISA Kits
- Apolipoprotein O ELISA Kits
- Apoptosis Inhibitor 5 ELISA Kits
- Apoptosis-Inducing Factor, Mitochondrion-Associated, 1 ELISA Kits
- Apotransferrin ELISA Kits
- APP ELISA Kits
- APPBP2 ELISA Kits
- Aprataxin ELISA Kits
- Aprotinin ELISA Kits
- APRT ELISA Kits



