TJP1 Antibodies
Your search for reliable TJP1 antibodies ends here. TJP1, known by aliases such as TJP1, Tjp1, LOC396567, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Pig, Cow, or other species, our range of TJP1 antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, FACS, IHC, ELISA, IF (p), providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Goat, Mouse. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our TJP1 antibodies in your research endeavors for dependable TJP1 detection.
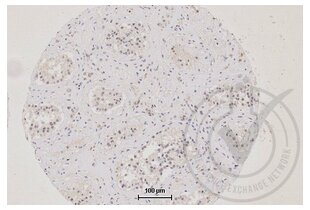
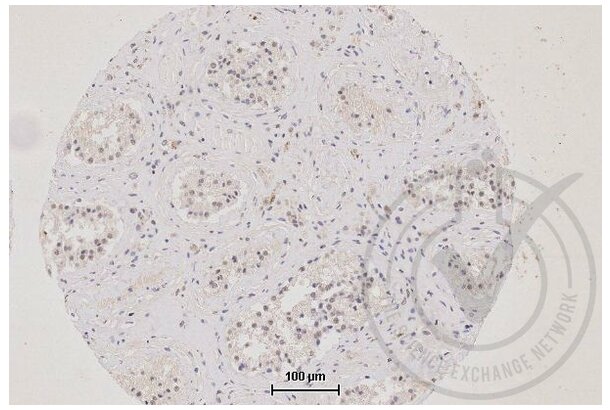 TJP1 antibody (AA 1551-1702) (ABIN675024)
TJP1 antibody (AA 1551-1702) (ABIN675024)
TJP1 Reactivity: Human, Mouse, Rat, Pig, Cow WB, ELISA, FACS, IHC (p), IF (p), IF (cc), IHC (fro) Host: Rabbit Polyclonal unconjugated
TJP1 Reactivity: Human, Mouse, Rat, Pig, Cow, Dog, Guinea Pig, Rabbit, Horse, Monkey, Chicken, Avian, Cat, Donkey, Goat, Hamster, Sheep WB, IF, IHC (p), IHC (fro) Host: Goat Polyclonal unconjugated
KO Validated TJP1 Reactivity: Human WB, IP, IF, ICC Host: Rabbit Polyclonal unconjugated
TJP1 Antibodies by Grade
Find TJP1 Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
TJP1 Antibodies by Host
Find TJP1 Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
TJP1 Antibodies by Clonality
Find available monoclonal or polyclonal TJP1 Antibodies. Click on a link to go to the corresponding products.
TJP1 Antibodies by Clone
Find TJP1 Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
Popular TJP1 Antibodies
- (7)
- (7)
- (1)
- (2)
- (11)
- (1)
- (8)
- (6)
- (7)
- (1)
- (6)
- (1)
- (6)
- (2)
- (3)
- (4)
- (4)
- (3)
- (1)
- (2)
- (2)
- (2)
- (2)
- (2)
Latest Publications for our TJP1 Antibodies
: "Epithelial Heat Shock Proteins Mediate the Protective Effects of Limosilactobacillus reuteri in Dextran Sulfate Sodium-Induced Colitis." in: Frontiers in immunology, Vol. 13, pp. 865982, (2022) (PubMed).: "Pinocembrin alleviates ulcerative colitis in mice via regulating gut microbiota, suppressing TLR4/MD2/NF-κB pathway and promoting intestinal barrier." in: Bioscience reports, Vol. 40, Issue 7, (2021) (PubMed).
: "3D Organoid Culture From Adult Salivary Gland Tissues as an ex vivo Modeling of Salivary Gland Morphogenesis." in: Frontiers in cell and developmental biology, Vol. 9, pp. 698292, (2021) (PubMed).
: "Parasite-Derived Excretory-Secretory Products Alleviate Gut Microbiota Dysbiosis and Improve Cognitive Impairment Induced by a High-Fat Diet." in: Frontiers in immunology, Vol. 12, pp. 710513, (2021) (PubMed).
: "Psychoactive Effects of Lactobacillus johnsonii Against Restraint Stress-Induced Memory Dysfunction in Mice Through Modulating Intestinal Inflammation and permeability-a Study Based on the Gut-Brain ..." in: Frontiers in pharmacology, Vol. 12, pp. 662148, (2021) (PubMed).
: "Early-Life Intervention Using Exogenous Fecal Microbiota Alleviates Gut Injury and Reduce Inflammation Caused by Weaning Stress in Piglets." in: Frontiers in microbiology, Vol. 12, pp. 671683, (2021) (PubMed).
: "The Antimicrobial Peptide Mastoparan X Protects Against Enterohemorrhagic Escherichia coli O157:H7 Infection, Inhibits Inflammation, and Enhances the Intestinal Epithelial Barrier." in: Frontiers in microbiology, Vol. 12, pp. 644887, (2021) (PubMed).
: "Astragalus membranaceus and Salvia miltiorrhiza ameliorates cyclosporin A-induced chronic nephrotoxicity through the "gut-kidney axis"." in: Journal of ethnopharmacology, Vol. 269, pp. 113768, (2021) (PubMed).
: "Supplemental Bacillus subtilis DSM 29784 and enzymes, alone or in combination, as alternatives for antibiotics to improve growth performance, digestive enzyme activity, anti-oxidative status, immune ..." in: The British journal of nutrition, Vol. 125, Issue 5, pp. 494-507, (2021) (PubMed).
: "Zizyphus jujuba cv. Muzao polysaccharides enhance intestinal barrier function and improve the survival of septic mice." in: Journal of food biochemistry, Vol. 45, Issue 5, pp. e13722, (2021) (PubMed).
Aliases for TJP1 Antibodies
tight junction protein 1 (TJP1) Antibodiestight junction protein 1 (Tjp1) Antibodies
tight junction protein ZO-1 (LOC396567) Antibodies
ZO-1 Antibodies
ZO1 Antibodies
zo1 Antibodies
Did you look for something else?
- TJAP1 Antibodies
- Titin Antibodies
- Tissue factor Antibodies
- TIRAP Antibodies
- TIPRL Antibodies
- TIPIN Antibodies
- Tiparp Antibodies
- TINF2 Antibodies
- TINAGL1 Antibodies
- TINAG Antibodies
- TIMP4 Antibodies
- TIMP3 Antibodies
- TIMP1 Antibodies
- TIMM9 Antibodies
- TIMM8B Antibodies
- TIMM8A/DDP Antibodies
- TIMM50 Antibodies
- TIMM44 Antibodies
- TIMM29 Antibodies
- TIMM23 Antibodies
- TJP2 Antibodies
- TJP3 Antibodies
- TK1 Antibodies
- TK2 Antibodies
- TKT Antibodies
- TKTL2 Antibodies
- TLE1 Antibodies
- TLE2 Antibodies
- TLE3 Antibodies
- TLE4 Antibodies
- TLE6 Antibodies
- TLK1 Antibodies
- TLK2 Antibodies
- TLL1 Antibodies
- TLN1 Antibodies
- TLN2 Antibodies
- TLR1 Antibodies
- TLR10 Antibodies
- TLR11 Antibodies
- TLR12 Antibodies



