MTOR Antibodies
Your search for reliable MTOR antibodies ends here. MTOR, known by aliases such as MTOR, Mtor, mtor, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Cow, Fish, or other species, our range of MTOR antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, ELISA, FACS, IF (cc), IF (p), providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Goat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our MTOR antibodies in your research endeavors for dependable MTOR detection.
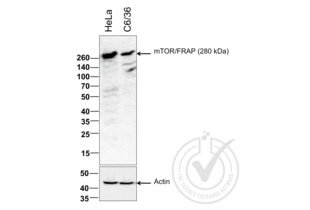
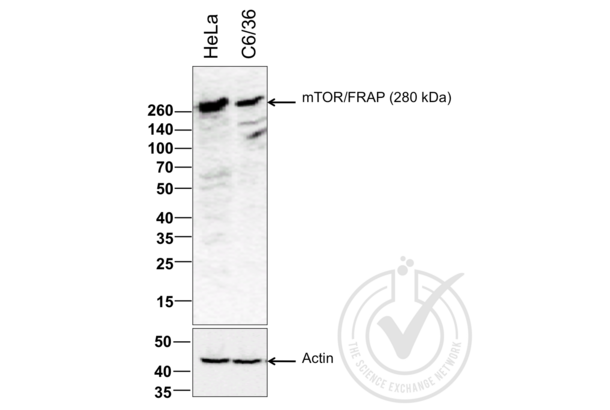 MTOR antibody (AA 2436-2492) (ABIN676403)
MTOR antibody (AA 2436-2492) (ABIN676403)
mTOR Reactivity: Human, Mouse, Rat WB, ELISA, FACS, IHC (p), IF (p), IF (cc), IHC (fro) Host: Rabbit Polyclonal unconjugated
mTOR Reactivity: Human, Mouse, Rat WB, IF, IHC (p), IP, ICC, ChIP Host: Rabbit Polyclonal unconjugated
mTOR Reactivity: Human WB, IHC, IP, ICC Host: Rabbit Polyclonal unconjugated
MTOR Antibodies by Grade
Find MTOR Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
MTOR Antibodies by Host
Find MTOR Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
MTOR Antibodies by Clonality
Find available monoclonal or polyclonal MTOR Antibodies. Click on a link to go to the corresponding products.
Popular MTOR Antibodies
- (9)
- (3)
- (1)
- (5)
- (15)
- (7)
- (7)
- (7)
- (4)
- (3)
- (4)
- (5)
- (5)
- (3)
- (5)
- (2)
- (3)
- (3)
- (1)
- (2)
- (3)
- (3)
Latest Publications for our MTOR Antibodies
: "Downregulation of the ubiquitin ligase KBTBD8 prevented epithelial ovarian cancer progression." in: Molecular medicine (Cambridge, Mass.), Vol. 26, Issue 1, pp. 96, (2021) (PubMed).: "Linc-ROR promotes the progression of breast cancer and decreases the sensitivity to rapamycin through miR-194-3p targeting MECP2." in: Molecular oncology, Vol. 14, Issue 9, pp. 2231-2250, (2021) (PubMed).
: "Cynaropicrin Induces Cell Cycle Arrest and Apoptosis by Inhibiting PKM2 to Cause DNA Damage and Mitochondrial Fission in A549 Cells." in: Journal of agricultural and food chemistry, Vol. 69, Issue 45, pp. 13557-13567, (2021) (PubMed).
: "Long non-coding RNA (lncRNA) H19 induces hepatic steatosis through activating MLXIPL and mTORC1 networks in hepatocytes." in: Journal of cellular and molecular medicine, Vol. 24, Issue 2, pp. 1399-1412, (2020) (PubMed).
: "ZNF322A-mediated protein phosphorylation induces autophagosome formation through modulation of IRS1-AKT glucose uptake and HSP-elicited UPR in lung cancer." in: Journal of biomedical science, Vol. 27, Issue 1, pp. 75, (2020) (PubMed).
: "HDAC6, modulated by miR-206, promotes endometrial cancer progression through the PTEN/AKT/mTOR pathway." in: Scientific reports, Vol. 10, Issue 1, pp. 3576, (2020) (PubMed).
: "Contribution of mTOR and PTEN to Radioresistance in Sporadic and NF2-Associated Vestibular Schwannomas: A Microarray and Pathway Analysis." in: Cancers, Vol. 12, Issue 1, (2020) (PubMed).
: "Metformin Protects Against Spinal Cord Injury by Regulating Autophagy via the mTOR Signaling Pathway." in: Neurochemical research, Vol. 43, Issue 5, pp. 1111-1117, (2019) (PubMed).
: "Pro-Inflammatory Activation of A New Immortalized Human Microglia Cell Line." in: Brain sciences, Vol. 9, Issue 5, (2019) (PubMed).
: "Phospho-mTOR expression in human glioblastoma microglia-macrophage cells." in: Neurochemistry international, Vol. 129, pp. 104485, (2019) (PubMed).
Aliases for MTOR Antibodies
mechanistic target of rapamycin kinase (MTOR) Antibodiesmechanistic target of rapamycin (MTOR) Antibodies
mechanistic target of rapamycin (serine/threonine kinase) (Mtor) Antibodies
mechanistic target of rapamycin kinase (Mtor) Antibodies
mechanistic target of rapamycin kinase (mtor) Antibodies
2610315D21Rik Antibodies
AI327068 Antibodies
flat Antibodies
FRAP Antibodies
FRAP1 Antibodies
Frap1 Antibodies
frap1 Antibodies
FRAP2 Antibodies
RAFT1 Antibodies
RAPT1 Antibodies
tor Antibodies
wu:fc22h08 Antibodies
Did you look for something else?
- MTMR9 Antibodies
- MTMR8 Antibodies
- MTMR4 Antibodies
- MTMR3 Antibodies
- MTMR2 Antibodies
- MTMR14 Antibodies
- MTMR12 Antibodies
- MTMR11 Antibodies
- MTMR10 Antibodies
- MTM1 Antibodies
- MTL5 Antibodies
- MTIF3 Antibodies
- MTIF2 Antibodies
- MTHFSD Antibodies
- MTHFS Antibodies
- MTHFR Antibodies
- MTHFD2L Antibodies
- MTHFD2 Antibodies
- MTHFD1L Antibodies
- MTHFD1 Antibodies
- MTPN Antibodies
- MTR Antibodies
- MTR4 Antibodies
- MTRF1 Antibodies
- MTRF1L Antibodies
- MTRNR2L1 Antibodies
- MTRR Antibodies
- MTSS1 Antibodies
- MTSS1L Antibodies
- MTTP Antibodies
- MTUS1 Antibodies
- MTX2 Antibodies
- MTX3 Antibodies
- Mu Opioid Receptor 1 Antibodies
- MUC1 Antibodies
- MUC13 Antibodies
- MUC15 Antibodies
- MUC16 Antibodies
- MUC19 Antibodies
- MUC2 Antibodies



