Reelin Antibodies
Your search for reliable Reelin antibodies ends here. Reelin, known by aliases such as RELN, Reln, reln, reln.L, LOC579060, is an integral part of our antibody range. Whether you're working with Human, Rat, Mouse, or other species, our range of Reelin antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, IHC, ELISA, ICC, IP, providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Goat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our Reelin antibodies in your research endeavors for dependable Reelin detection.
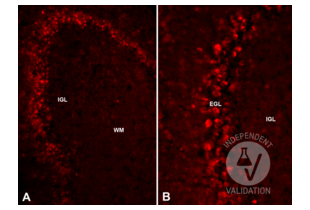
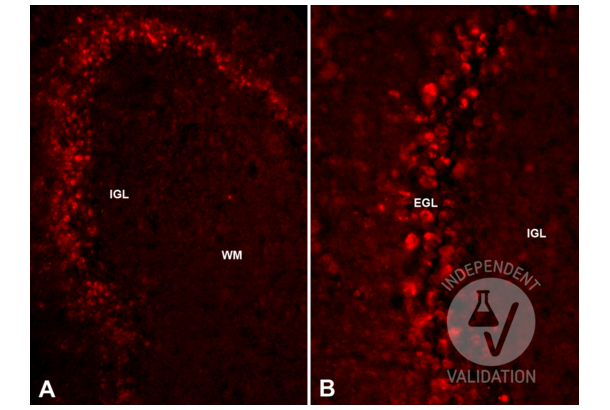 Reelin antibody (AA 3143-3461) (ABIN7435136)
Reelin antibody (AA 3143-3461) (ABIN7435136)
RELN Reactivity: Mouse WB, IHC, IP, ICC Host: Rabbit Polyclonal unconjugated
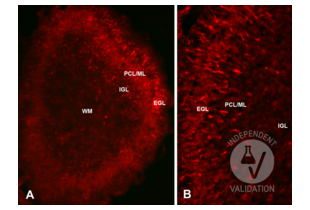
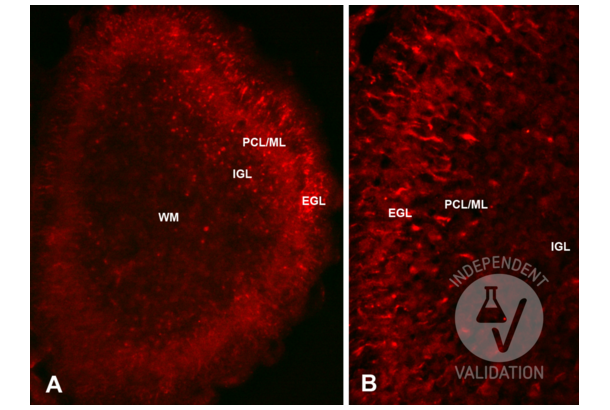 Reelin antibody (N-Term) (ABIN6264693)
Reelin antibody (N-Term) (ABIN6264693)
RELN Reactivity: Human, Rat, Mouse WB, IHC, ELISA Host: Rabbit Polyclonal unconjugated
RELN Reactivity: Human WB, IHC, IP, ICC Host: Mouse Monoclonal C8 unconjugated
Reelin Antibodies by Grade
Find Reelin Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
Reelin Antibodies by Reactivity
Find Reelin Antibodies for a variety of species such as anti-Human Reelin, anti-Rat Reelin, anti-Mouse Reelin. The species listed below are among those available. Click on a link to go to the corresponding products.
Reelin Antibodies by Application
Find Reelin Antibodies validated for a specific application such as WB, IHC, ELISA, ICC. Some of the available applications are listed below. Click on a link to go to the corresponding products.
Reelin Antibodies by Host
Find Reelin Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
Reelin Antibodies by Clonality
Find available monoclonal or polyclonal Reelin Antibodies. Click on a link to go to the corresponding products.
Reelin Antibodies by Clone
Find Reelin Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
Reelin Antibodies by Conjugate
Find Reelin Antibodies with a specific conjugate such as Biotin, FITC, Alexa Fluor 488. The conjugates listed below are among those available. Click on a link to go to the corresponding products.
Popular Reelin Antibodies
- (7)
- (1)
- (2)
- (1)
- (3)
- (3)
- (2)
- (2)
- (3)
- (3)
- (3)
- (3)
- (1)
- (1)
- (1)
- (1)
- (1)
Latest Publications for our Reelin Antibodies
: "Yin Yang 1 sustains biosynthetic demands during brain development in a stage-specific manner." in: Nature communications, Vol. 10, Issue 1, pp. 2192, (2019) (PubMed).: "Immunohistochemistry of aberrant neuronal development induced by 6-propyl-2-thiouracil in rats." in: Toxicology letters, Vol. 261, pp. 59-71, (2016) (PubMed).
: "Reelin expression is up-regulated in mice colon in response to acute colitis and provides resistance against colitis." in: Biochimica et biophysica acta, Vol. 1863, Issue 2, pp. 462-473, (2016) (PubMed).
: "Smad4 and Trim33/Tif1γ redundantly regulate neural stem cells in the developing cortex." in: Cerebral cortex (New York, N.Y. : 1991), Vol. 24, Issue 11, pp. 2951-63, (2014) (PubMed).
: "Glycidol induces axonopathy and aberrations of hippocampal neurogenesis affecting late-stage differentiation by exposure to rats in a framework of 28-day toxicity study." in: Toxicology letters, Vol. 224, Issue 3, pp. 424-32, (2014) (PubMed).
: "Reversible effect of maternal exposure to chlorpyrifos on the intermediate granule cell progenitors in the hippocampal dentate gyrus of rat offspring." in: Reproductive toxicology (Elmsford, N.Y.), Vol. 35, pp. 125-36, (2013) (PubMed).
: "Reversible effect of developmental exposure to chlorpyrifos on late-stage neurogenesis in the hippocampal dentate gyrus in mouse offspring." in: Reproductive toxicology (Elmsford, N.Y.), Vol. 38, pp. 25-36, (2013) (PubMed).
: "Glycidol induces axonopathy by adult-stage exposure and aberration of hippocampal neurogenesis affecting late-stage differentiation by developmental exposure in rats." in: Toxicological sciences : an official journal of the Society of Toxicology, Vol. 134, Issue 1, pp. 140-54, (2013) (PubMed).
: "Reversible aberration of neurogenesis targeting late-stage progenitor cells in the hippocampal dentate gyrus of rat offspring after maternal exposure to acrylamide." in: Archives of toxicology, Vol. 86, Issue 5, pp. 779-90, (2012) (PubMed).
: "Transient aberration of neuronal development in the hippocampal dentate gyrus after developmental exposure to brominated flame retardants in rats." in: Archives of toxicology, Vol. 86, Issue 9, pp. 1431-42, (2012) (PubMed).
Aliases for Reelin Antibodies
reelin (RELN) Antibodiesreelin (Reln) Antibodies
reelin (reln) Antibodies
reelin L homeolog (reln.L) Antibodies
reelin (LOC579060) Antibodies
LIS2 Antibodies
MGC172539 Antibodies
PRO1598 Antibodies
Reelen Antibodies
reeler Antibodies
reln Antibodies
RELN Antibodies
RL Antibodies
rl Antibodies
Rl Antibodies
Did you look for something else?
- RED1 Antibodies
- RECQL5 Antibodies
- RECQL4 Antibodies
- RECQL2 Antibodies
- RecQ Protein-Like (DNA Helicase Q1-Like) Antibodies
- Recoverin Antibodies
- RECK Antibodies
- Receptor Accessory Protein 4 Antibodies
- Receptor Accessory Protein 1 Antibodies
- REC8 Antibodies
- RDM1 Antibodies
- RDH8 Antibodies
- RDH16 Antibodies
- RDH13 Antibodies
- RDH12 Antibodies
- RDH11 Antibodies
- RDH10 Antibodies
- RDBP Antibodies
- RD3 Antibodies
- RCSD1 Antibodies
- REEP2 Antibodies
- REEP3 Antibodies
- REEP5 Antibodies
- REEP6 Antibodies
- REG1 Antibodies
- REG1A Antibodies
- REG1B Antibodies
- REG3A Antibodies
- REG3B Antibodies
- REG3g Antibodies
- REG4 Antibodies
- Regucalcin Antibodies
- Regulating Synaptic Membrane Exocytosis 1 Antibodies
- Regulator of G Protein Signaling 9 Binding Protein Antibodies
- Regulator of G-Protein Signaling 5 Antibodies
- Relaxin Antibodies
- Relaxin 1 Antibodies
- Relaxin 2 Antibodies
- Relaxin 3 Antibodies
- Relaxin 3 Receptor 1 Antibodies



