Glucagon Antibodies
Glucagon also has other effects on metabolism. It promotes the breakdown of fats in adipose tissue, leading to the release of fatty acids into the bloodstream. It can also stimulate the production of ketone bodies, which are alternative energy sources derived from fat metabolism. Additionally, glucagon can inhibit the synthesis of glycogen and fatty acids, further contributing to the mobilization of stored energy.
Overall, the role of glucagon is to maintain glucose homeostasis in the body by increasing blood glucose levels when they are low. It works in coordination with insulin to regulate blood sugar levels and ensure that they stay within a normal range.
The main applications of anti-Glucagon antibodies include:
- Immunohistochemistry (IHC): Glucagon antibodies are used in IHC to detect and visualize Glucagon in tissue samples, particularly in pancreatic tissue sections.
- Immunocytochemistry (ICC): These Glucagon antibodies are also used in ICC to detect and localize Glucagon in cultured cells.
- Western Blot: Glucagon antibodies can be used in Western blotting techniques to detect Glucagon.
- ELISA: Enzyme-Linked Immunosorbent Assay (ELISA) can be performed using anti-Glucagon antibodies to quantify the levels of Glucagon in biological samples.
- Flow Cytometry: Glucagon antibodies can be used in flow cytometry (FACS) to analyze and sort cells based on their Glucagon expression levels.
GCG Reactivity: Human WB, IF, IHC (p), FACS Host: Rabbit Polyclonal RB19525 unconjugated
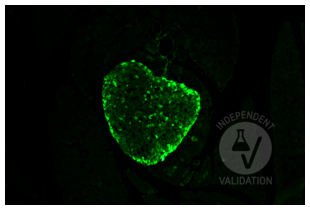
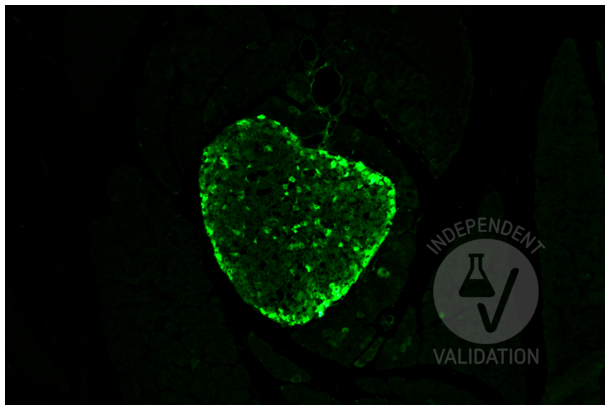 Glucagon antibody (ABIN7448121)
Glucagon antibody (ABIN7448121)
GCG Reactivity: Human ELISA, WB, IHC, IF, FM Host: Rabbit Polyclonal unconjugated
GCG Reactivity: Mouse WB, IHC, IP, ICC Host: Rabbit Polyclonal unconjugated
Glucagon Antibodies by Host
Find Glucagon Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
Glucagon Antibodies by Clonality
Find available monoclonal or polyclonal Glucagon Antibodies. Click on a link to go to the corresponding products.
Popular Glucagon Antibodies
- (6)
- (2)
- (1)
- (5)
- (4)
- (3)
- (3)
- (3)
- (6)
- (3)
- (2)
- (1)
- (1)
- (1)
- (1)
- (2)
- (2)
- (3)
- (3)
Latest Publications for our Glucagon Antibodies
: "Emodin Alleviates Severe Acute Pancreatitis-Associated Acute Lung Injury by Inhibiting the Cold-Inducible RNA-Binding Protein (CIRP)-Mediated Activation of the NLRP3/IL-1β/CXCL1 Signaling." in: Frontiers in pharmacology, Vol. 12, pp. 655372, (2021) (PubMed).: "HWL-088, a new potent free fatty acid receptor 1 (FFAR1) agonist, improves glucolipid metabolism and acts additively with metformin in ob/ob diabetic mice." in: British journal of pharmacology, Vol. 177, Issue 10, pp. 2286-2302, (2021) (PubMed).
: "Identification of Potential Therapeutic Targets and Pathways of Liraglutide Against Type 2 Diabetes Mellitus (T2DM) Based on Long Non-Coding RNA (lncRNA) Sequencing." in: Medical science monitor : international medical journal of experimental and clinical research, Vol. 26, pp. e922210, (2020) (PubMed).
: "Necrostatin-1 supplementation enhances young porcine islet maturation and in vitro function." in: Xenotransplantation, pp. e12555, (2019) (PubMed).
: "Chronic oscillating glucose challenges disarrange innate immune homeostasis to potentiate the variation of neutrophil-lymphocyte ratio in rats with or without hidden diabetes mellitus." in: Diabetes, metabolic syndrome and obesity : targets and therapy, Vol. 11, pp. 277-288, (2018) (PubMed).
: "Abnormal neutrophil signature in the blood and pancreas of presymptomatic and symptomatic type 1 diabetes. ..." in: JCI insight, Vol. 3, Issue 18, (2018) (PubMed).
: "The Polycomb-Dependent Epigenome Controls β Cell Dysfunction, Dedifferentiation, and Diabetes." in: Cell metabolism, Vol. 27, Issue 6, pp. 1294-1308.e7, (2018) (PubMed).
: "Antidiabetic drug glyburide modulates depressive-like behavior comorbid with insulin resistance." in: Journal of neuroinflammation, Vol. 14, Issue 1, pp. 210, (2018) (PubMed).
: "Low dose doxycycline decreases systemic inflammation and improves glycemic control, lipid profiles, and islet morphology and function in db/db mice." in: Scientific reports, Vol. 7, Issue 1, pp. 14707, (2017) (PubMed).
: "Activin Enhances α- to β-Cell Transdifferentiation as a Source For β-Cells In Male FSTL3 Knockout Mice." in: Endocrinology, Vol. 157, Issue 3, pp. 1043-54, (2016) (PubMed).
Aliases for Glucagon Antibodies
glucagon (GCG) Antibodiesglucagon (Gcg) Antibodies
glucagon L homeolog (gcg.L) Antibodies
GCG Antibodies
gcg-A Antibodies
gcg1 Antibodies
GLP-1 Antibodies
GLP1 Antibodies
GLP2 Antibodies
Glu Antibodies
GRPP Antibodies
PPG Antibodies
Did you look for something else?
- GLTP Antibodies
- GLT8D2 Antibodies
- GLT8D1 Antibodies
- GLT6D1 Antibodies
- GLT25D2 Antibodies
- GLS2 Antibodies
- GLRX5 Antibodies
- GLRX3 Antibodies
- GLRB Antibodies
- GLRA4 Antibodies
- GLRA3 Antibodies
- GLRa2 Antibodies
- GLRA1 Antibodies
- GLP2R Antibodies
- GLP1R Antibodies
- GLP-2 Antibodies
- GLP-1 Antibodies
- Glomulin, FKBP Associated Protein Antibodies
- GLOD4 Antibodies
- GLO1 Antibodies
- Glucagon Receptor Antibodies
- Glucocorticoid Receptor Antibodies
- Glucocorticoid Receptor beta Antibodies
- Glucose-6-Phosphate Dehydrogenase Antibodies
- Glucuronic Acid Epimerase Antibodies
- Glucuronidase beta Antibodies
- GLUD2 Antibodies
- GLUT1 Antibodies
- GLUT12 Antibodies
- GLUT4 Antibodies
- Glutamate Dehydrogenase 1 Antibodies
- Glutamate Receptor 1 Antibodies
- Glutamate Receptor 3 Antibodies
- Glutamate Receptor, Ionotropic, AMPA 4 Antibodies
- Glutamate Receptor, Ionotropic, Kainate 5 Antibodies
- Glutamate-Ammonia Ligase Antibodies
- Glutamic Pyruvate Transaminase (Alanine Aminotransferase) 2 Antibodies
- Glutamic-Oxaloacetic Transaminase 2, Mitochondrial (Aspartate Aminotransferase 2) Antibodies
- Glutaminase Antibodies
- Glutaredoxin 1 Antibodies



