E-cadherin Antibodies
Your search for reliable E-cadherin antibodies ends here. E-cadherin, known by aliases such as CDH1, si:busm1-180o5.2, cdh1, Cdh1, cdh1.S, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Monkey, Pig, or other species, our range of E-cadherin antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, IHC, FACS, ELISA, IF, providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Goat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our E-cadherin antibodies in your research endeavors for dependable E-cadherin detection.
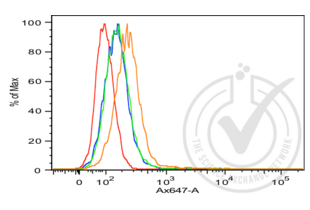
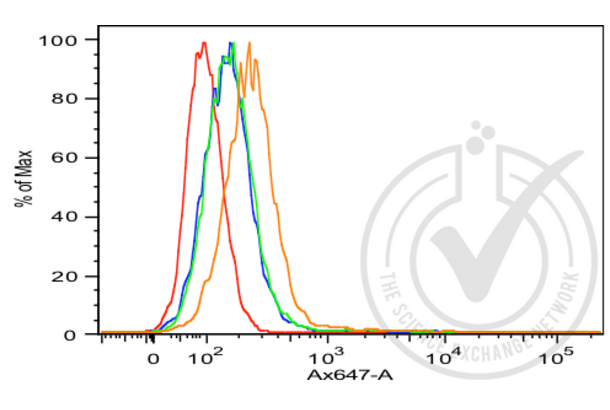 E-cadherin antibody (AA 401-500) (ABIN1387847)
E-cadherin antibody (AA 401-500) (ABIN1387847)
CDH1 Reactivity: Human, Mouse, Rat WB, FACS, ELISA, IHC (p), ICC, IF (p), IHC (fro), IF (cc) Host: Rabbit Polyclonal unconjugated
KO Validated CDH1 Reactivity: Human WB, IF, IP, IHC (p), ICC, IHC (wm) Host: Rabbit Polyclonal unconjugated
CDH1 Reactivity: Human, Mouse, Rat WB, IHC, ELISA, IF, ICC Host: Rabbit Polyclonal unconjugated
E-cadherin Antibodies by Grade
Find E-cadherin Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
E-cadherin Antibodies by Host
Find E-cadherin Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
E-cadherin Antibodies by Clonality
Find available monoclonal or polyclonal E-cadherin Antibodies. Click on a link to go to the corresponding products.
E-cadherin Antibodies by Fragment
Find E-cadherin Antibodies with a specific Fragment. The Fragment listed below are among those available. Click on a link to go to the corresponding products.
Popular E-cadherin Antibodies
- (14)
- (12)
- (1)
- (10)
- (8)
- (19)
- (5)
- (1)
- (6)
- (6)
- (2)
- (5)
- (9)
- (2)
- (2)
- (1)
- (5)
- (5)
- (5)
- (5)
- (5)
- (30)
- (3)
- (1)
- (5)
Latest Publications for our E-cadherin Antibodies
: "Characterization of the prognostic values and response to immunotherapy/chemotherapy of Krüppel-like factors in prostate cancer." in: Journal of cellular and molecular medicine, Vol. 24, Issue 10, pp. 5797-5810, (2021) (PubMed).: "38" in: , Vol. 1363, Issue Nucleic acids research, pp. 28-39, (1991)
: "Loss of RE-1 silencing transcription factor accelerates exocrine damage from pancreatic injury." in: Cell death & disease, Vol. 11, Issue 2, pp. 138, (2020) (PubMed).
: "BST2 regulates interferon gamma-dependent decrease in invasion of HTR-8/SVneo cells via STAT1 and AKT signaling pathways and expression of E-cadherin." in: Cell adhesion & migration, Vol. 14, Issue 1, pp. 24-41, (2020) (PubMed).
: "Low Dose of Paclitaxel Combined with XAV939 Attenuates Metastasis, Angiogenesis and Growth in Breast Cancer by Suppressing Wnt Signaling." in: Cells, Vol. 8, Issue 8, (2020) (PubMed).
: "Host restriction of Escherichia coli recurrent urinary tract infection occurs in a bacterial strain-specific manner." in: PLoS pathogens, Vol. 14, Issue 12, pp. e1007457, (2019) (PubMed).
: "Deletion of Nrf2 enhances susceptibility to eosinophilic sinonasal inflammation in a murine model of rhinosinusitis." in: International forum of allergy & rhinology, Vol. 9, Issue 1, pp. 114-119, (2019) (PubMed).
: "Nickel exposure induces persistent mesenchymal phenotype in human lung epithelial cells through epigenetic activation of ZEB1." in: Molecular carcinogenesis, Vol. 57, Issue 6, pp. 794-806, (2019) (PubMed).
: "Loss of BCL9/9l suppresses Wnt driven tumourigenesis in models that recapitulate human cancer." in: Nature communications, Vol. 10, Issue 1, pp. 723, (2019) (PubMed).
: "Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p ..." in: Journal of experimental & clinical cancer research : CR, Vol. 38, Issue 1, pp. 62, (2019) (PubMed).
Aliases for E-cadherin Antibodies
cadherin 1 (CDH1) Antibodiessi:busm1-180o5.2 (si:busm1-180o5.2) Antibodies
cadherin 1, type 1, E-cadherin (epithelial) (cdh1) Antibodies
cadherin 1 (Cdh1) Antibodies
cadherin 1, type 1 S homeolog (cdh1.S) Antibodies
AA960649 Antibodies
Arc-1 Antibodies
arc-1 Antibodies
c-cadherin Antibodies
cadherin-1 Antibodies
CD324 Antibodies
cd324 Antibodies
cdh1 Antibodies
cdhc-A Antibodies
CDHE Antibodies
cdhe Antibodies
E-Cad Antibodies
E-cad Antibodies
E-cadherin Antibodies
ECAD Antibodies
ecad Antibodies
Ecad Antibodies
l-cam Antibodies
L-CAM Antibodies
LCAM Antibodies
lcam Antibodies
si:dz180o5.2 Antibodies
Um Antibodies
UVO Antibodies
uvo Antibodies
uvomorulin Antibodies
xb-cad Antibodies
XB-cadherin Antibodies
XBcad Antibodies
xcdh1 Antibodies
XTCAD-1 Antibodies
Did you look for something else?
- DZIP3 Antibodies
- DZIP1 Antibodies
- DZANK1 Antibodies
- DYX1C1 Antibodies
- Dystrophin Antibodies
- Dystroglycan Antibodies
- Dystrobrevin Binding Protein 1 Antibodies
- Dystrobrevin beta Antibodies
- Dystonin Antibodies
- Dysferlin Antibodies
- DYRK4 Antibodies
- DYRK3 Antibodies
- DYRK2 Antibodies
- DYRK1B Antibodies
- DYRK1A Antibodies
- DYNLT3 Antibodies
- DYNLT1 Antibodies
- DYNLRB2 Antibodies
- DYNLRB1 Antibodies
- DYNLL2 Antibodies
- E2F1 Antibodies
- E2F2 Antibodies
- E2F3 Antibodies
- E2F4 Antibodies
- E2F4/E2F5 Antibodies
- E2F5 Antibodies
- E2F6 Antibodies
- E2F7 Antibodies
- E2F8 Antibodies
- E4F1 Antibodies
- EAF1 Antibodies
- EAF2 Antibodies
- Early Growth Response 2 Antibodies
- EARS2 Antibodies
- EBF1 Antibodies
- EBF2 Antibodies
- EBF3 Antibodies
- EBF4 Antibodies
- EBI3 Antibodies
- EBLN2 Antibodies



