NOTCH2 Antibodies
Your search for reliable NOTCH2 antibodies ends here. NOTCH2, known by aliases such as LOC581186, NOTCH2, notch3, Notch2, notch2, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Pig, Dog, or other species, our range of NOTCH2 antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, ELISA, IF (cc), IF (p), FACS, providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Rat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our NOTCH2 antibodies in your research endeavors for dependable NOTCH2 detection.
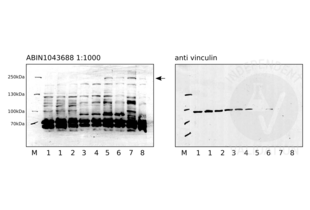
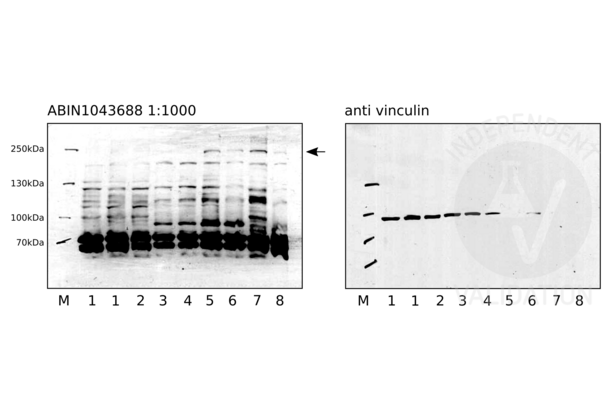 NOTCH2 antibody (ABIN104899)
NOTCH2 antibody (ABIN104899)
NOTCH2 Reactivity: Human WB, ELISA, IHC Host: Rabbit Polyclonal unconjugated
NOTCH2 Reactivity: Human, Mouse, Rat WB, ELISA, IHC Host: Rabbit Polyclonal unconjugated
NOTCH2 Reactivity: Human, Mouse WB, ChIP-seq Host: Rabbit Polyclonal unconjugated
NOTCH2 Antibodies by Grade
Find NOTCH2 Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
NOTCH2 Antibodies by Reactivity
Find NOTCH2 Antibodies for a variety of species such as anti-Human NOTCH2, anti-Mouse NOTCH2, anti-Rat NOTCH2. The species listed below are among those available. Click on a link to go to the corresponding products.
NOTCH2 Antibodies by Host
Find NOTCH2 Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
NOTCH2 Antibodies by Clonality
Find available monoclonal or polyclonal NOTCH2 Antibodies. Click on a link to go to the corresponding products.
NOTCH2 Antibodies by Clone
Find NOTCH2 Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
Popular NOTCH2 Antibodies
- (3)
- (1)
- (1)
- (3)
- (1)
- (2)
- (2)
- (2)
- (2)
- (2)
- (2)
- (1)
- (1)
- (3)
- (2)
- (1)
- (1)
- (1)
Latest Publications for our NOTCH2 Antibodies
: "Potassium depletion induces cellular conversion in the outer medullary collecting duct altering Notch signaling pathway." in: Scientific reports, Vol. 10, Issue 1, pp. 5708, (2020) (PubMed).: "Protein Expression Analysis in Uterine Cervical Cancer for Potential Targets in Treatment." in: Pathology oncology research : POR, (2018) (PubMed).
: "Epistatic Association of CD14 and NOTCH2 Genetic Polymorphisms with Biliary Atresia in a Southern Chinese Population." in: Molecular therapy. Nucleic acids, Vol. 13, pp. 590-595, (2018) (PubMed).
: "RNA granule assembly and disassembly modulated by nuclear factor associated with double-stranded RNA 2 and nuclear factor 45." in: The Journal of biological chemistry, Vol. 289, Issue 30, pp. 21163-80, (2015) (PubMed).
: "Fatty acid binding protein 4 deficiency protects against oxygen-induced retinopathy in mice." in: PLoS ONE, Vol. 9, Issue 5, pp. e96253, (2015) (PubMed).
: "Interference of Notch 2 inhibits the progression of gliomas and induces cell apoptosis by induction of the cell cycle at the G0/G1 phase." in: Molecular medicine reports, Vol. 11, Issue 1, pp. 734-8, (2014) (PubMed).
: "Lysophosphatidic acid increases proximal tubule cell secretion of profibrotic cytokines PDGF-B and CTGF through LPA2- and G?q-mediated Rho and ?v?6 integrin-dependent activation of TGF-?." in: The American journal of pathology, Vol. 181, Issue 4, pp. 1236-49, (2012) (PubMed).
: "ADAM10 is essential for Notch2-dependent marginal zone B cell development and CD23 cleavage in vivo." in: The Journal of experimental medicine, Vol. 207, Issue 3, pp. 623-35, (2010) (PubMed).
: "Immunolocalization of notch signaling protein molecules in a maxillary chondrosarcoma and its recurrent tumor." in: European journal of medical research, Vol. 15, Issue 10, pp. 456-60, (2010) (PubMed).
: "Notch4 overexpression in ameloblastoma correlates with the solid/multicystic phenotype." in: Oral surgery, oral medicine, oral pathology, oral radiology, and endodontics, Vol. 110, Issue 2, pp. 224-33, (2010) (PubMed).
Aliases for NOTCH2 Antibodies
neurogenic locus notch homolog protein 2 (LOC581186) Antibodiesnotch 2 (NOTCH2) Antibodies
notch 3 (notch3) Antibodies
notch 2 (Notch2) Antibodies
notch 2 (notch2) Antibodies
AGS2 Antibodies
AI853703 Antibodies
HJCYS Antibodies
hN2 Antibodies
N2 Antibodies
notch6 Antibodies
sb:cb884 Antibodies
Did you look for something else?
- Notch1 Antibodies
- NOTCH-Regulated Ankyrin Repeat Protein Antibodies
- NOSTRIN Antibodies
- NOSIP Antibodies
- NOS2 Antibodies
- NOS1AP Antibodies
- NOS1 Antibodies
- Norrie Disease (Pseudoglioma) Antibodies
- Noradrenaline Antibodies
- NOP58 Antibodies
- NOP56 Antibodies
- NOP2 Antibodies
- NOP16 Antibodies
- Nop14 Antibodies
- Nop10 Antibodies
- NONO Antibodies
- NOMO2 Antibodies
- NOMO1 Antibodies
- NOM1 Antibodies
- NOLC1 Antibodies
- NOTCH2NL Antibodies
- NOTCH3 Antibodies
- NOTCH4 Antibodies
- NOTO Antibodies
- NOTUM Antibodies
- NOV Antibodies
- NOVA1 Antibodies
- NOVA2 Antibodies
- NOX1 Antibodies
- NOX3 Antibodies
- NOX5 Antibodies
- NOXA Antibodies
- NOXA1 Antibodies
- NOXO1 Antibodies
- NPAP1 Antibodies
- NPAS1 Antibodies
- NPAS2 Antibodies
- NPAS3 Antibodies
- NPAS4 Antibodies
- NPAT Antibodies



