CTCF Antibodies
Your search for reliable CTCF antibodies ends here. CTCF, known by aliases such as CTCF, ctcf, Ctcf, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Cow, Dog, or other species, our range of CTCF antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, IHC, ELISA, IF, IP, providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Rat. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our CTCF antibodies in your research endeavors for dependable CTCF detection.
CTCF Reactivity: Human WB, IHC, IF, ChIP, IP Host: Rabbit Polyclonal unconjugated

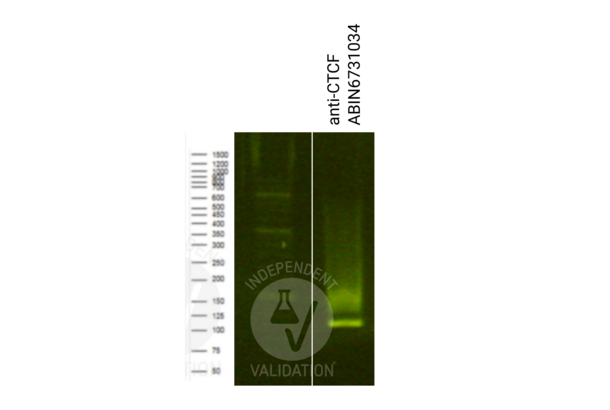 Recombinant CTCF antibody (ABIN6731034)
Recombinant CTCF antibody (ABIN6731034)
CTCF Reactivity: Human WB, ChIP-seq, CUT&RUN Host: Rabbit Monoclonal unconjugated Recombinant Antibody
CTCF Reactivity: Human, Mouse, Rat, Monkey IHC, IF Host: Rabbit Polyclonal unconjugated
CTCF Antibodies by Grade
Find CTCF Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
CTCF Antibodies by Host
Find CTCF Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
CTCF Antibodies by Clonality
Find available monoclonal or polyclonal CTCF Antibodies. Click on a link to go to the corresponding products.
CTCF Antibodies by Clone
Find CTCF Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
CTCF Antibodies by Conjugate
Find CTCF Antibodies with a specific conjugate such as Alexa Fluor 488, Alexa Fluor 555, Alexa Fluor 594. The conjugates listed below are among those available. Click on a link to go to the corresponding products.
Popular CTCF Antibodies
- (6)
- (2)
- (1)
- (5)
- (5)
- (4)
- (3)
- (3)
- (3)
- (3)
- (4)
- (2)
- (3)
- (1)
- (2)
- (2)
- (3)
Latest Publications for our CTCF Antibodies
: "The Human GATA1 Gene Retains a 5' Insulator That Maintains Chromosomal Architecture and GATA1 Expression Levels in Splenic Erythroblasts." in: Molecular and cellular biology, Vol. 35, Issue 10, pp. 1825-37, (2015) (PubMed).: "Domain-Specific and Stage-Intrinsic Changes in Tcrb Conformation during Thymocyte Development." in: Journal of immunology (Baltimore, Md. : 1950), Vol. 195, Issue 3, pp. 1262-72, (2015) (PubMed).
: "CTCF binding to the first intron of the major immediate early (MIE) gene of human cytomegalovirus (HCMV) negatively regulates MIE gene expression and HCMV replication." in: Journal of virology, Vol. 88, Issue 13, pp. 7389-401, (2014) (PubMed).
: "DNA methylation prevents CTCF-mediated silencing of the oncogene BCL6 in B cell lymphomas." in: The Journal of experimental medicine, Vol. 207, Issue 9, pp. 1939-50, (2010) (PubMed).
: "Loss of imprinting of IGF2 and H19, loss of heterozygosity of IGF2R and CTCF, and Helicobacter pylori infection in laryngeal squamous cell carcinoma." in: Journal of molecular medicine (Berlin, Germany), Vol. 86, Issue 9, pp. 1057-66, (2008) (PubMed).
: "The insulator factor CTCF controls MHC class II gene expression and is required for the formation of long-distance chromatin interactions." in: The Journal of experimental medicine, Vol. 205, Issue 4, pp. 785-98, (2008) (PubMed).
: "Cohesins localize with CTCF at the KSHV latency control region and at cellular c-myc and H19/Igf2 insulators." in: The EMBO journal, Vol. 27, Issue 4, pp. 654-66, (2008) (PubMed).
: "The protein CTCF is required for the enhancer blocking activity of vertebrate insulators." in: Cell, Vol. 98, Issue 3, pp. 387-96, (1999) (PubMed).
: "A widely expressed transcription factor with multiple DNA sequence specificity, CTCF, is localized at chromosome segment 16q22.1 within one of the smallest regions of overlap for common deletions in breast and prostate cancers." in: Genes, chromosomes & cancer, Vol. 22, Issue 1, pp. 26-36, (1998) (PubMed).
: "An exceptionally conserved transcriptional repressor, CTCF, employs different combinations of zinc fingers to bind diverged promoter sequences of avian and mammalian c-myc oncogenes." in: Molecular and cellular biology, Vol. 16, Issue 6, pp. 2802-13, (1996) (PubMed).
Aliases for CTCF Antibodies
CCCTC-binding factor (CTCF) AntibodiesCCCTC-binding factor (zinc finger protein) (ctcf) Antibodies
CCCTC-binding factor (ctcf) Antibodies
CCCTC-binding factor (Ctcf) Antibodies
AW108038 Antibodies
CTCF Antibodies
im:7152666 Antibodies
xctcf Antibodies
Did you look for something else?
- CTC1 Antibodies
- CTBS Antibodies
- CTBP2 Antibodies
- CTBP1 Antibodies
- CTAGE4 Antibodies
- CTAGE1 Antibodies
- CTAG2 Antibodies
- CTAG1B Antibodies
- CTAG1A Antibodies
- CT47A1 Antibodies
- CSTL1 Antibodies
- CSTF3 Antibodies
- CSTF2T Antibodies
- CSTF2 Antibodies
- CSTF1 Antibodies
- CSTB Antibodies
- CSTA Antibodies
- CST9L Antibodies
- CST9 Antibodies
- CST8 Antibodies
- CTCFL Antibodies
- CTD (Carboxy-Terminal Domain, RNA Polymerase II, Polypeptide A) Small Phosphatase Like 2 Antibodies
- CTDNEP1A Antibodies
- CTDP1 Antibodies
- CTDSP1 Antibodies
- CTDSP2 Antibodies
- CTDSPL Antibodies
- CTF18 Antibodies
- CTGF Antibodies
- CTH Antibodies
- CTHRC1 Antibodies
- CTIF/KIAA0427 Antibodies
- CTLA4 Antibodies
- CTNNA1 Antibodies
- CTNNA2 Antibodies
- CTNNA3 Antibodies
- CTNNAL1 Antibodies
- CTNNB1 Antibodies
- CTNNBIP1 Antibodies
- CTNNBL1 Antibodies



