Myc Proto-Oncogene protein Antibodies
Your search for reliable c-MYC antibodies ends here. c-MYC, known by aliases such as MYC, LOC100136746, Nol3, Myc, myca, myc.L, myc.S, mycb, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Dog, Tag, or other species, our range of c-MYC antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, ELISA, IHC, FACS, IF, providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse, Chicken. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our c-MYC antibodies in your research endeavors for dependable c-MYC detection.
MYC Reactivity: Human WB, ELISA, IF, IHC (p) Host: Mouse Monoclonal 1G7 unconjugated
MYC Reactivity: Human, Mouse WB, IF, IP, ICC, ChIP Host: Rabbit Polyclonal unconjugated
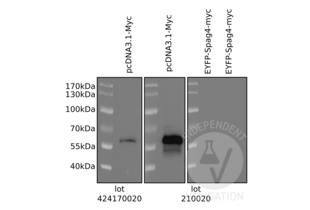
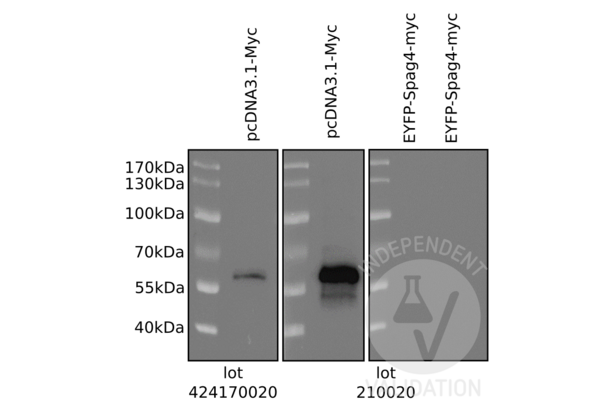 c-MYC antibody (AA 31-80) (ABIN1532205)
c-MYC antibody (AA 31-80) (ABIN1532205)
MYC Reactivity: Human, Mouse, Rat WB, IHC, ELISA, IF Host: Rabbit Polyclonal unconjugated
Myc Proto-Oncogene protein Antibodies by Grade
Find Myc Proto-Oncogene protein Antibodies with a specific Grade. The Grade listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein Antibodies by Reactivity
Find Myc Proto-Oncogene protein Antibodies for a variety of species such as anti-Human Myc Proto-Oncogene protein, anti-Mouse Myc Proto-Oncogene protein, anti-Rat Myc Proto-Oncogene protein. The species listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein Antibodies by Host
Find Myc Proto-Oncogene protein Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
Myc Proto-Oncogene protein Antibodies by Clonality
Find available monoclonal or polyclonal Myc Proto-Oncogene protein Antibodies. Click on a link to go to the corresponding products.
Popular Myc Proto-Oncogene protein Antibodies
- (1)
- (6)
- (5)
- (8)
- (3)
- (2)
- (12)
- (12)
- (7)
- (6)
- (2)
- (3)
- (4)
- (3)
- (3)
- (3)
- (1)
- (2)
- (3)
- (13)
- (2)
- (2)
Latest Publications for our Myc Proto-Oncogene protein Antibodies
: "Oxidative stress-induced mitophagy is suppressed by the miR-106b-93-25 cluster in a protective manner." in: Cell death & disease, Vol. 12, Issue 2, pp. 209, (2021) (PubMed).: "Long non-coding RNA CTSLP8 mediates ovarian cancer progression and chemotherapy resistance by modulating cellular glycolysis and regulating c-Myc expression through PKM2." in: Cell biology and toxicology, (2021) (PubMed).
: "Wnt/β‑catenin signaling modulates piperine‑mediated antitumor effects on human osteosarcoma cells." in: Molecular medicine reports, Vol. 21, Issue 5, pp. 2202-2208, (2021) (PubMed).
: "Manipulating ZmEXPA4 expression ameliorates the drought-induced prolonged anthesis and silking interval in maize." in: The Plant cell, Vol. 33, Issue 6, pp. 2058-2071, (2021) (PubMed).
: "ACSL4 promotes hepatocellular carcinoma progression via c-Myc stability mediated by ERK/FBW7/c-Myc axis." in: Oncogenesis, Vol. 9, Issue 4, pp. 42, (2020) (PubMed).
: "Cellular density-dependent increases in HIF-1α compete with c-Myc to down-regulate human EP4 receptor promoter activity through Sp-1-binding region." in: Pharmacology research & perspectives, Vol. 6, Issue 6, pp. e00441, (2019) (PubMed).
: "Neoisoliquiritigenin Inhibits Tumor Progression by Targeting GRP78-β- catenin Signaling in Breast Cancer." in: Current cancer drug targets, Vol. 18, Issue 4, pp. 390-399, (2018) (PubMed).
: "Sphingomyelin synthase 2 overexpression promotes cisplatin-induced apoptosis of HepG2 cells." in: Oncology letters, Vol. 15, Issue 1, pp. 483-488, (2018) (PubMed).
: "Upregulation of E‑cadherin expression mediated by a novel dsRNA suppresses the growth and metastasis of bladder cancer cells by inhibiting β-catenin/TCF target genes." in: International journal of oncology, Vol. 52, Issue 6, pp. 1815-1826, (2018) (PubMed).
: "Functional characterization of chloroplast-targeted RbgA GTPase in higher plants." in: Plant molecular biology, Vol. 95, Issue 4-5, pp. 463-479, (2018) (PubMed).
Aliases for Myc Proto-Oncogene protein Antibodies
MYC proto-oncogene, bHLH transcription factor (MYC) Antibodiesv-myc myelocytomatosis viral oncogene homolog (LOC100136746) Antibodies
nucleolar protein 3 (apoptosis repressor with CARD domain) (Nol3) Antibodies
MYC proto-oncogene, bHLH transcription factor (Myc) Antibodies
myelocytomatosis oncogene (Myc) Antibodies
MYC proto-oncogene, bHLH transcription factor a (myca) Antibodies
MYC proto-oncogene, bHLH transcription factor L homeolog (myc.L) Antibodies
MYC proto-oncogene, bHLH transcription factor S homeolog (myc.S) Antibodies
v-myc myelocytomatosis viral oncogene homolog (avian) (MYC) Antibodies
MYC proto-oncogene, bHLH transcription factor b (mycb) Antibodies
CG10798 gene product from transcript CG10798-RB (Myc) Antibodies
anon-WO03040301.171 Antibodies
ARC Antibodies
AU016757 Antibodies
B430311C09Rik Antibodies
bHLHe39 Antibodies
bHLHe57 Antibodies
c-Myc Antibodies
c-myc Antibodies
C-MYC Antibodies
c-MYC Antibodies
c-myc II Antibodies
CG10798 Antibodies
CMYC Antibodies
cmyc Antibodies
CMyc Antibodies
D-Myc Antibodies
d-myc Antibodies
da Antibodies
DM Antibodies
Dm Antibodies
dm Antibodies
dm/dMyc Antibodies
dm/myc Antibodies
Dmel\CG10798 Antibodies
DMYc Antibodies
Dmyc Antibodies
dMYC Antibodies
dMyc Antibodies
dmyc Antibodies
dMyc1 Antibodies
dmyc1 Antibodies
EG:BACN5I9.1 Antibodies
l(1)G0139 Antibodies
l(1)G0354 Antibodies
l(1)G0359 Antibodies
LOC100136746 Antibodies
mMyc Antibodies
MRTL Antibodies
MYC Antibodies
myc Antibodies
myc-B Antibodies
myc1 Antibodies
Myc2 Antibodies
myc2 Antibodies
MYCC Antibodies
Niard Antibodies
Nird Antibodies
NOP Antibodies
Nop30 Antibodies
RNCMYC Antibodies
zc-myc Antibodies
zgc:55680 Antibodies
Did you look for something else?
- MYBPHL Antibodies
- MYBPH Antibodies
- MYBPC3 Antibodies
- MYBPC2 Antibodies
- MYBPC1 Antibodies
- MYBL2 Antibodies
- MYBL1 Antibodies
- MYBBP1A Antibodies
- Myb-Related Transcription Factor, Partner of Profilin Antibodies
- MYB Antibodies
- MYADM Antibodies
- MXI1 Antibodies
- MXD4 Antibodies
- MXD3 Antibodies
- MXD1 Antibodies
- MX2 Antibodies
- MX1 Antibodies
- MVP Antibodies
- MVK Antibodies
- MVD Antibodies
- Myc Target 1 Antibodies
- MYCBP Antibodies
- MYCBP2 Antibodies
- MYCBPAP Antibodies
- MYCN Antibodies
- Mycobacterium Tuberculosis Antibodies
- MYDGF Antibodies
- MYEF2 Antibodies
- Myeloid Differentiation Primary Response Gene (88) Antibodies
- Myeloperoxidase Antibodies
- MYF5 Antibodies
- MYF6 Antibodies
- MYH1 Antibodies
- MYH10 Antibodies
- MYH11 Antibodies
- MYH13 Antibodies
- MYH15 Antibodies
- MYH2 Antibodies
- MYH3 Antibodies
- MYH4 Antibodies



