Elastin Antibodies
Your search for reliable Elastin antibodies ends here. Elastin, known by aliases such as ELN, Eln, LOC574132, Tcur_2481, Kfla_2289, LOC100620140, is an integral part of our antibody range. Whether you're working with Human, Mouse, Rat, Dog, Guinea Pig, or other species, our range of Elastin antibodies offer precise detection across diverse samples. These specialized antibodies are tailored for various scientific applications like WB, IHC, ELISA, IP, IHC (p), providing you with options like polyclonal, recombinant, and monoclonal antibodies, sourced from different host species such as Rabbit, Mouse. The efficacy of our antibodies is well-established, demonstrated through multiple methods.
Detailed information, including references, images, and validations by other customers, can be found on each product page. Should you require assistance in finding a specific product, our customer service team is ready to assist. Utilize our Elastin antibodies in your research endeavors for dependable Elastin detection.
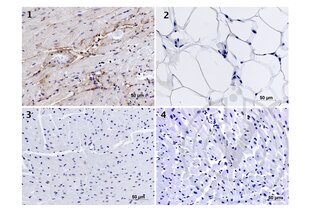
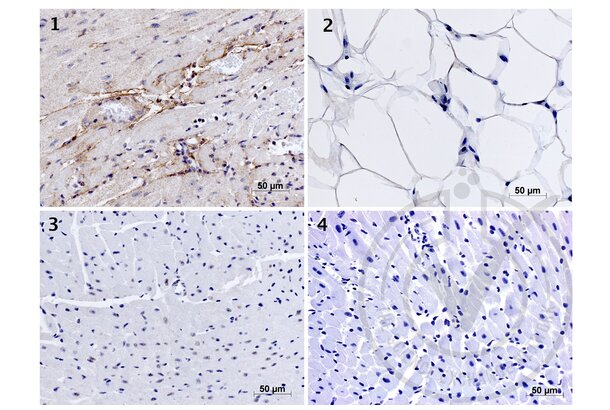 Elastin antibody (AA 681-786) (ABIN734003)
Elastin antibody (AA 681-786) (ABIN734003)
ELN Reactivity: Human, Mouse, Rat WB, ELISA, IHC (p), IHC (fro), IF (cc), IF (p), FACS Host: Rabbit Polyclonal unconjugated
ELN Reactivity: Human WB, ELISA, ICC, IHC (p), IHC (fro) Host: Rabbit Polyclonal unconjugated
ELN Reactivity: Mouse WB, IHC, IP, ICC Host: Rabbit Polyclonal unconjugated
Elastin Antibodies by Reactivity
Find Elastin Antibodies for a variety of species such as anti-Human Elastin, anti-Mouse Elastin, anti-Rat Elastin. The species listed below are among those available. Click on a link to go to the corresponding products.
Elastin Antibodies by Host
Find Elastin Antibodies with a specific Host. The Host listed below are among those available. Click on a link to go to the corresponding products.
Elastin Antibodies by Binding Specificity
Find Elastin Antibodies with a specific epitope. The epitopes listed below are among those available. Click on a link to go to the corresponding products.
Elastin Antibodies by Clonality
Find available monoclonal or polyclonal Elastin Antibodies. Click on a link to go to the corresponding products.
Elastin Antibodies by Clone
Find Elastin Antibodies with a specific Clone. The Clone listed below are among those available. Click on a link to go to the corresponding products.
Elastin Antibodies by Conjugate
Find Elastin Antibodies with a specific conjugate such as Biotin, FITC, Alexa Fluor 488. The conjugates listed below are among those available. Click on a link to go to the corresponding products.
Popular Elastin Antibodies
- (4)
- (7)
- (1)
- (8)
- (3)
- (3)
- (3)
- (3)
- (3)
- (2)
- (2)
- (2)
- (1)
- (1)
- (1)
- (2)
Latest Publications for our Elastin Antibodies
: "Tropoelastin: A novel marker for plaque progression and instability." in: Circulation. Cardiovascular imaging, Vol. 11, Issue 8, (2018) (PubMed).: "Vasoprotective effects of urocortin 1 against atherosclerosis in vitro and in vivo." in: PLoS ONE, Vol. 9, Issue 12, pp. e110866, (2014) (PubMed).
: "Stimulatory effects of cardiotrophin 1 on atherosclerosis." in: Hypertension, Vol. 62, Issue 5, pp. 942-50, (2013) (PubMed).
: "Elastogenesis at the onset of human cardiac valve development." in: Development (Cambridge, England), Vol. 140, Issue 11, pp. 2345-53, (2013) (PubMed).
: "Immunofluorescence and fluorescent-protein tagging show high correlation for protein localization in mammalian cells." in: Nature methods, Vol. 10, Issue 4, pp. 315-23, (2013) (PubMed).
: "Piezoelectric PU/PVDF electrospun scaffolds for wound healing applications." in: Colloids and surfaces. B, Biointerfaces, Vol. 96, pp. 29-36, (2012) (PubMed).
: "The influence of elastic components of the venous wall on the biomechanical properties of different veins used for arterial reconstruction." in: European journal of vascular and endovascular surgery : the official journal of the European Society for Vascular Surgery, Vol. 40, Issue 2, pp. 224-9, (2010) (PubMed).
: "Impact of cryopreservation on extracellular matrix structures of heart valve leaflets." in: The Annals of thoracic surgery, Vol. 81, Issue 3, pp. 918-26, (2006) (PubMed).
: "Connection between elastin haploinsufficiency and increased cell proliferation in patients with supravalvular aortic stenosis and Williams-Beuren syndrome." in: American journal of human genetics, Vol. 71, Issue 1, pp. 30-44, (2002) (PubMed).
: "Characterization of biologically active domains on elastin: identification of a monoclonal antibody to a cell recognition site." in: Biochemistry, Vol. 25, Issue 18, pp. 5172-6, (1986) (PubMed).
Aliases for Elastin Antibodies
elastin (ELN) Antibodieselastin (Eln) Antibodies
elastin (LOC574132) Antibodies
elastin (Tcur_2481) Antibodies
elastin (Kfla_2289) Antibodies
elastin (LOC100620140) Antibodies
AI385707 Antibodies
AI480567 Antibodies
E030024M20Rik Antibodies
ELN Antibodies
RATTREL11 Antibodies
SVAS Antibodies
TREL11 Antibodies
Trela Antibodies
Trela26 Antibodies
WBS Antibodies
WS Antibodies
Did you look for something else?
- Elastase 3B Antibodies
- Elastase Antibodies
- ELAPOR1 Antibodies
- ELANE Antibodies
- ELAC2 Antibodies
- ELAC1 Antibodies
- EIF6 Antibodies
- EIF5B Antibodies
- EIF5AL1 Antibodies
- EIF5A2 Antibodies
- EIF5 Antibodies
- EIF4H Antibodies
- EIF4G3 Antibodies
- EIF4G2 Antibodies
- EIF4G1 Antibodies
- EIF4ENIF1 Antibodies
- EIF4EBP3 Antibodies
- eIF4EBP2 Antibodies
- eIF4EBP1 Antibodies
- EIF4E2 Antibodies
- ELAV (Embryonic Lethal, Abnormal Vision, Drosophila)-Like 1 (Hu Antigen R) Antibodies
- ELAVL2 Antibodies
- ELAVL4 Antibodies
- ELF1 Antibodies
- ELF2 Antibodies
- ELF3 Antibodies
- Elf4 Antibodies
- ELF5 Antibodies
- ELFN1 Antibodies
- ELFN2 Antibodies
- ELK1 Antibodies
- ELK3 Antibodies
- ELK4 Antibodies
- ELL Antibodies
- ELL2 Antibodies
- ELL3 Antibodies
- ELMO1 Antibodies
- ELMO2 Antibodies
- ELMO3 Antibodies
- ELMOD1 Antibodies



